

-
水资源短缺是一个全球性环境问题,影响着数亿人的正常生产生活. 再生水因其可靠、经济等优点,成为城市非常规水源而被广泛应用. 数据显示,在美国加利福尼亚州,较早就有约25%再生水作为各类湖泊生态用水的补充;在日本大阪,大约50%的再生水被用于城市景观水景. 随着“海绵城市”建设的全国推进,以污水处理厂尾水或再生水为补给水源的城市湿地公园近年来在国内也大量涌现,在生态生境、城市景观、游憩活动等多种生态经济服务中发挥着重要的作用. 数据显示中国约有50%再生水用于补充城市水体,约13.2%城市水景受再生水补充[1]. 但是,由于再生水中COD、总氮和总磷等污染物通常较高(高于《地表水环境质量标准GB3838-2002》中限值),使受再生水补给的水体面临富营养化污染问题.
为减轻再生水带来的污染,需要采取水污染控制技术来削减或控制污染. 其中,底泥疏浚可以消除内源性污染,但工程量较大,经济成本较高[2]. 絮凝剂等化学药剂的投加,可以去除氮磷等营养物,但易导致二次污染[3]. 水生植物修复方法不但能够通过植物和微生物代谢作用转化去除水中污染物,还有助于重新构建的水生生态系统平衡,促进氮、磷等物质的长期循环,具有成本低、环境友好、兼具景观效果等特点,因而受到广泛应用[4]. 虽然水生植物对氮、磷和有机物等具有良好的去除能力,但是修复效果往往受到许多外界条件的影响和限制[5]. 例如,水生植物生长的最佳水温为15—28 ℃,过高的水温不利于水生植物向水体复氧,从而使夏季表现出较差的修复效果[6]. 另外,pH值、光照条件和种植密度等都会影响水生植物修复性能. 现阶段,大多数对植物修复效能的研究均在实验室模拟装置内进行,很难对实际场景中的植物修复措施进行复现. 而再生水的水质特征是其有机污染物多为难降解有机物,氮磷浓度偏高,碳源不足. 对于以再生水作为补给水源的水体,对实际应用中的效果以及季节变化等影响并不了解. 水生植物能否在城市水体应用中起到良好的恢复水质的效果仍然不太清楚.
圆明园是一个著名的遗址公园,由于北京所面临的缺水问题,该园自2007年以来一直以再生水作为唯一的补水水源,年补水量约900万m3[7]. 为净化水质,园区持续多年采取了水生植物修复措施. 本研究选择圆明园作为对象,研究了再生水进入园区后,水体中COD、TP和TN等污染物的时空分布与变化,结合水体理化性质和水生植物生长状况,分析了水生植物对水体的修复效果与机制,以期为再生水的实际应用提供理论支撑.
-
圆明园地处北纬40°00′08′'至40°00′41′'、东经116°16′59′' to 116°18′08′'之间,属半湿润季风气候. 年均降雨量约630 mm,其中约80%集中在夏季. 圆明园长期以再生水作为唯一水源,且园区没有出水口,水体相对封闭,能在干扰较小的情况下研究再生水对其水体的影响. 现阶段,园区已采取一定的水质修复措施,包括特定区域的水生植物种植,秋冬季不定期的植物残体打捞,以及避免周边土地的化肥使用等. 同时,区域内的植物种类、密度呈现出明显的空间差异,有利于对比分析水生植物在城市水体中的实际修复效能. 因此,选取该区域作为代表性对象开展研究. 研究区内水生植物包括:芦苇(Phragmites australis)、香蒲(Typha orientalis,)、富贵莲(Nelumbo sp. )、菹草(Potamogeton crispus)、水盾草(Cabomba caroliniana A. Gray)和苦草(Vallisneria natans)等. 这些植物均为多年生植物,能够在水体中长期存活. 其中,富贵莲、苦草、水盾草、菹草是园区主要水生植物,其生物量和覆盖面积超过所有水生植物的90%. 如图1所示,根据水生植物覆盖度的不同,分为植物修复区域(覆盖度≥80%)、半修复区域(覆盖度40%—80%)和非植物修复区域(覆盖度≤40%). 在不同类型的植物种植区域均匀布设共43个采样点,将进水点(1#)和重点游览景区等均涵盖在内. 其中,1—3、6、8、10、14—20、25、28—30、42、43采样点共19个点属于植物修复区域;7、9、21、27、37采样点共5个属于半修复区域;4、5、11—13、22—24、26、31—36、38—41采样点共19个属于非植物修复区域. 园区内总体水流方向呈现为自西北向东南.
-
除1、2月水面结冰无法采样外,2022年3月—12月每月采集表层水样. 利用GPS定位仪准确定位各采样点位置. 使用有机玻璃采水器与水面下20 cm处采集表层水样品. 样品采集后,在现场使用便携式多功能水质分析仪(SL1000,哈希)测定温度、溶解氧(DO)、氧化还原电位(ORP)和pH值. 水样于4 ℃条件下避光运输及保存,48 h内完成测试. 使用快速消解分光光度法(HJ/T 399-2007)、碱性过硫酸钾消解紫外分光光度法(HJ 636-2012)、钼酸铵分光光度法(GB
11893 -89)测定COD、TN和TP浓度. 所有数据均为3次重复实验平均值. -
使用ArcGIS 10.5软件(Esri,California,美国)和1:
6500 的研究区地图,在WGS 1984坐标系下运行. 应用基于协方差函数的普通克里格插值和ArcGIS的扩展模块制作各类污染物的空间分布图[8]. 如式(1)所示,使用半变异函数模型对污染物浓度进行拟合:式中:γ(h)为已知点xi和xj的半变异;h为这两点间的距离;z为属性值.
-
圆明园水体测试的理化指标主要是水温、DO、pH和ORP值,能够反映水体环境的基本物理化学性质. 如图2所示,圆明园水体水温随季节变化明显,春夏秋冬四季的平均水温分别是18.0、29.5、20.2、3.3 ℃.
但季节对水体pH值影响较小,四季pH平均值在6.6—8.1之间,相比于1号入水口的6.3—7.3,再生水进入后,水体pH值呈总体上升趋势,这是由于水生植物光合作用消耗了CO2和碳酸盐而导致[9]. 水体冬季DO平均含量最高(12.78 mg∙L−1),夏季(6.00 mg∙L−1)和秋季(4.51 mg∙L−1)则较低. 夏季高温使水体中微生物活性增加,以及秋季水生植物的腐败,都消耗了水中大量的溶解氧[10]. 氧化还原电位的季节变化趋势与DO相似,呈现冬季(230 mV)高,夏季(144 mV)低的特征. 水体DO和ORP值在夏秋季偏低的状况,意味着水体在夏秋季面临更高的水质改善需要.
-
由于受再生水处理工艺的影响,1号点再水生入口处的总氮浓度较高,全年处于6.19—11.58 mg∙L−1之间(如图3),尤其是冬季污水处理中反硝化效率偏低[11],导致来水TN可高达11.58 mg∙L−1. 尽管如此,距出水口最远的37号点,TN可降至2.22—3.72 mg∙L−1,表明流经过程所设置的水生植物修复区域发挥了很大的去除作用.
图3还显示,不同的路径处表现出了不同的TN水平. 24和32号点分别是福海两个流量较大的入湖口,再生水经北部河段(11、22—24号点)和南部河段(7—9号点、28—32号点)流入福海,但是这两个河段的TN却呈现很大差别. 春夏两季中,TN在北部河段始终处于较高水平;但在南部河段8号点处却明显下降,并一直保持在相对较低水平,且此种影响一直延续到福海入口处. 与1号点相比,32号点的TN全年均值削减了57.9%,而24号点处仅为23.5%. 北部河段水生植物覆盖度不足5%,而南部河段8号点和28—30号点周围均有大量水生植物生长(图1). 结果说明,水生植物对再生水中的含氮污染物具有非常好的净化处理能力,且不受低碳水平的影响.
水温对硝化菌和反硝化菌的除氮效能影响很大[12],这导致研究区域内TN存在着明显的季节差异,秋、冬季的2.70—11.54 mg∙L−1和2.29—11.58 mg∙L−1明显高于春、夏季的0.39—7.10 mg∙L−1和0.46—6.27 mg∙L−1(图3). 中游植物修复区域(6、8、14—21、28—30号点)春夏两季的平均TN浓度分别为2.90 mg∙L−1和1.84 mg∙L−1,也显著低于秋(5.07 mg∙L−1)、冬(4.48 mg∙L−1)两季. 春夏季旺盛的水生植物生长,使TN在春夏季中游区域的削减率分别达到58.4%和70.3%,水质处于地表Ⅳ—Ⅴ间. 受秋冬季TN来水偏高的影响,中游区域TN在秋冬季偏高,但是秋冬季TN削减率也达到56.1%和61.4%. 再生水中总氮主要以硝态氮为主,水生植物可以通过根、茎、叶直接从水中吸收含氮物质[13],但硝态氮往往难以直接被植物吸收[14]. 因此,相比于水生植物直接吸收同化的除氮过程,叶片所附着微生物的代谢过程是氮去除的主要途径[15]. 一方面,沉水植物茂密的叶片为微生物提供了附着基质和栖息场所,有利于形成附着生物膜. 其中含有大量的氨化细菌和硝化细菌,他们的丰度远高于缺少植物的水体或底泥[16]. 水中的有机氮被氨化细菌转化为铵态氮,又经过硝化和反硝化,最终转化为N2排出水体[17]. 另一方面,水生植物根际周围吸附了红薇菌属等具有高固氮能力的微生物, 并在nifH、 nifD和nifK等固氮功能基因介导下提升了根际周围沉积物的固氮能力[18]. 同时,植物的代谢还能提供充足的有机质和氧气,进一步促进微生物的生长硝化反应的发生. 秋冬季水温较低,植物生长减慢,碳源相对不足,微生物活性降低. 在这种情况下,硝化和反硝化的能力被减弱,因而降低了氮的转化分解能力[19]. 即使如此,大量沉水植物种植,仍然实现了秋冬季TN的大量去除,有效保障再生水补给区域内TN的削减.
-
磷是引起藻华的关键元素,因此磷的控制对城市水体的生态管理尤为重要. 与TN的季节变化不同,TP平均浓度在夏季最高而冬季最低,分别为0.132 mg∙L−1和0.047 mg∙L−1. 1号入水口处的TP浓度在夏季达到了0.162 mg∙L−1,而冬季仅为0.096 mg∙L−1,较高的磷入水使1号点及其附近区域在各季节均显示出较高的TP浓度. 对园区水体而言,水中磷不仅来源于再生水,还有很大一部分来源于水体的内源释放. 夏季相对较高的温度增加了假单胞菌等溶磷细菌的活性,促使了沉积物中的生物活性磷转化为可溶性的正磷酸盐,向水中释放[20]. 夏季再生水TP偏高和内源释放导致了园区水体TP水平的升高.
从图4还可以看出,TP在福海北部(11、22—24号点)和南部(7—9号点、28—32号点)两个河段中表现了与TN类似的空间变化规律. 北部河段的TP浓度较南部河段偏高,表明了植物修复对TP有良好的去除效果. 但TP进入福海后又明显升高,并未像TN一样在福海南部保持持续的低浓度. 这是由氮和磷不同的来源和去除机制所造成.
南部河段的水生植物通过茎和叶直接吸收和同化水中的可溶性磷酸,并通过主动运输将磷转移到根系和周围的沉积物中,转化成有机磷储存起来. 研究表明,植物丰富区域的底泥中总磷和有机磷含量显著高于其他区域[20]. 植物周边附着的浮游生物、单细胞绿藻和原生动物也能够分泌细胞外酶如碱性磷酸酶和磷酸二酯酶,从而降解水中的磷[21]. 福海及其周围游客密集,并有大量的观光游船在此往来. 游船螺旋桨频繁扰动湖底沉积物,大大增加了水体中颗粒态磷含量[7]. 水生植物茎叶能有效吸附悬浮颗粒物,并防止颗粒物再悬浮[5]. 但是,福海内水生植物种植稀少,使得游船搅起的颗粒物难以沉降,从而导致TP在福海内的再度升高.
为了解不同区域植物修复效果的差异,本研究选取了14—15号、18—21号、28—30号三个面积较大、植物覆盖程度较高的区域进行对比. 如表1所示,相比于1号点,三个区域在春、秋季都获得较高的TP去除率,可达到78.3%、63.4%和57.2%. 但是在夏季,各区域对TP的去除效率均有所降低,14—15号点和28—30号点分别下降到了34.4%和34.6%,但18—21号点却从秋季的63.4%大幅下降到只有6.6%. 由图1b可知,14—15号点和28—30号点区域内超半数是挺水植物富贵莲,而18—21号点内则以沉水植物菹草、苦草和水盾草为主,富贵莲只占较小一部分. 相比于沉水植物,挺水植物具有更发达的根系,对沉积物颗粒的固定作用也更强[22]. 另外,体型高大的挺水植物能够有效阻挡岸边形成的径流,减少陆源磷向水体中的输入. 夏季温度高且是雨季,径流和沉积物释放是夏季TP的两个重要来源. 因此,挺水植物区中的总磷去除效率在夏季所受到的影响较小.
-
如图5所示,四个季节COD浓度均沿水流方向呈现出西北低而东南高的趋势. 在春夏秋冬四个季节,研究区域水体COD的最高浓度分别达到了56.8、51.0、47.7、37.4 mg∙L−1,均高于地表Ⅲ类限值(20 mg∙L−1). 但是在西北端1号点的再生水入水处,其COD浓度在8.1—18.1 mg∙L−1之间,符合地表Ⅲ类水标准,说明再生水并非导致水体COD升高的直接原因. 随着水流不断流入,水体中的COD水平呈持续上升趋势. 同时,全年COD浓度呈先上升后下降的趋势,与水温变化趋势一致. 研究表明圆明园沉积物中的微生物以好氧微生物为主[20]. 在较高的温度下,沉积物与水柱之间的界面处有机质交换更为活跃[23],这一点可以被夏季显著降低的溶解氧证实(图2c). 并且,随着水体流动距离的增加,其承接了越来越多来自沉积物释放的有机污染物. 下游区域水流速度缓慢,释放的有机物质无法被即使稀释,更加重了该区域的COD污染. 因此,上述时空分布趋势表明有机物的释放和颗粒的再悬浮是造成园区COD超标的主要原因之一.
从空间分布图可以看出,全年高COD浓度的区域较为一致,主要位于后湖西南(4—8号点)和福海东岸(35—38、42—43号点). 其四季平均COD浓度分别为41.4、41.9、41.5、26.6 mg∙L−1,且最高达到56.8 mg∙L−1. 这些区域是研究区域内游客活动较为密集的地方. 沿岸行人的频繁走动,将陆地的土壤和灰尘扬起,通过空气沉降进入邻近水体[24]. 同时,福海水面观光游船的往来搅动了沉积物,使底质颗粒再悬浮进入水中,增加了水中颗粒态有机物的浓度. 更重要的是,这些区域水生植物覆盖率偏低,难以起到固定周围沉积物颗粒以及吸附水体有机物的作用[25],这使得COD在这些区域内难以被去除而不断累积. 植物的缺失也无法为沉积物中的好氧微生物提供充足的氧气,使其无法进行正常的有机物代谢,因此大量有机物被释放到水体中. 相对应地,14—15号点、18—21号点、28—30号的3个典型的植物修复区域中全年COD平均浓度分别为17.7—32.9、26.1—38.0和25.1—32.2 mg∙L−1. 尽管这些区域的COD浓度均高于入水点处,但相较于非植物修复区域仍有明显的下降. 水生植物可以直接吸收水中有机物质并将其用于自身的细胞合成和代谢活动[4]. 同时,其叶片表面是好氧细菌、微藻以及原生动物的良好栖息地,它们形成的生物微膜能够高效降解水中的有机物[15]. 此外,植物光合作用提高了水中DO浓度,提高了有机污染物的降解效率. 尽管水生植物对COD有一定的修复效果,但其控制作用相较氮、磷污染而言比较弱. 总的来看,植物修复在城市公园水体中具有良好的应用前景,但其效果受季节、植物种类等影响较大. 建议在实际应用时考虑各种类型植物的合理配比,并在污染严重的季节采取例如投加药剂、底泥覆盖等方法强化水质恢复.
-
1)研究区域TN、TP、COD的季节平均浓度分别为1.84—5.07、0.047—0.132、26.6—41.9 mg∙L−1. 除冬季的TP外,其余所有季节的3个污染指标均高于地表Ⅲ类水标准.
2)再生水输入显著地影响了TN、TP的浓度,是园区内TN、TP的主要来源,但其并非是造成COD浓度超标的主要原因.
3)水生植物修复总体对TN表现出良好的去除效果,但受季节影响较大. 在温度较高的春、夏季,水生植物对TN的修复效果较好,但对TP的修复效能却随着温度上升而出现明显下降.
4)挺水植物对TP有着更加出色稳定的修复效能,但对TN和COD的修复效能与沉水植物之间相差不大.
水生植物修复改善再生水补给水体水质效能的研究
Effect of aquatic plants on water quality restoration in the urban water bodies recharged by reclaimed water
-
摘要: 再生水已成为缺水地区的重要非常规水源. 但是,再生水中的营养性污染物质往往高于地表水水质要求,易导致受纳水体出现富营养化污染. 本文选取了一个仅以再生水为水源补充的城市水体进行研究. 根据水生植物生长状况,划分为修复程度不同的区域,使用普通克里金插值法获取了再生水流入后的总氮(TN)、总磷(TP)、化学需氧量(COD)等污染物的空间分布与变化,分析了水生植物修复的效能与机制. 结果显示,水生植物能够在再生水流入下游之前削减57.9%的TN,且在春、夏季修复效果更加突出. 植物修复区域最高削减了78.3%的TP,挺水植物区对TP的修复效果较为稳定. 但受高温影响,沉水植物区对TP的削减率从最高的63.4%下降到了夏季的6.61%. 由于沉积物的持续释放,植物修复对COD的去除效率比对TN和TP低,仅在沉水植物区表现了相对较好的修复效果.Abstract: Reclaimed water (RW) has become an important unconventional water source in water-scarce areas. However, the nutrient pollutants in RW are higher than the limits in the standard of surface water quality, which lead to the eutrophication pollution in the waterbody recharged by RW. In this study, an urban waterbody using RW as the sole water source was selected as a case. The waters were divided into areas with different phytoremediation degrees according to the growth status of aquatic plants. The spatial and temporal variations of pollutants including total nitrogen (TN), total phosphorus (TP) and chemical oxygen demand (COD) after the inflow of RW were obtained by using the common Kriging interpolation method. The efficiency and mechanism of phytoremediation were analyzed. The results showed that the replenishment of RW was a crucial reason for TN and TP pollution. Aquatic plants reduced 57.9% of TN at the highest level before the RW flows downstream, and the remediation effect was more prominent in spring and summer. The highest reduction rate of TP in phytoremediation area was 78.3%. The reduction rate of TP in the area with submerged plant decreased from the highest 63.4% to 6.61% in summer due to the high temperature, while that of emergent plant on TP was relatively stable. Due to the continuous release of sediment, the removal efficiency of COD was lower than those of TN and TP, and the remediation effect was relatively outstanding only in submerged plant areas.
-
-

图 1 采样点布设、水流方向和圆明园游客活动路线(a)以及植物修复状况(b)
Figure 1. Sampling point locations, water flow direction, and tourism activities situation (a), and areas of phytoremediation (b)
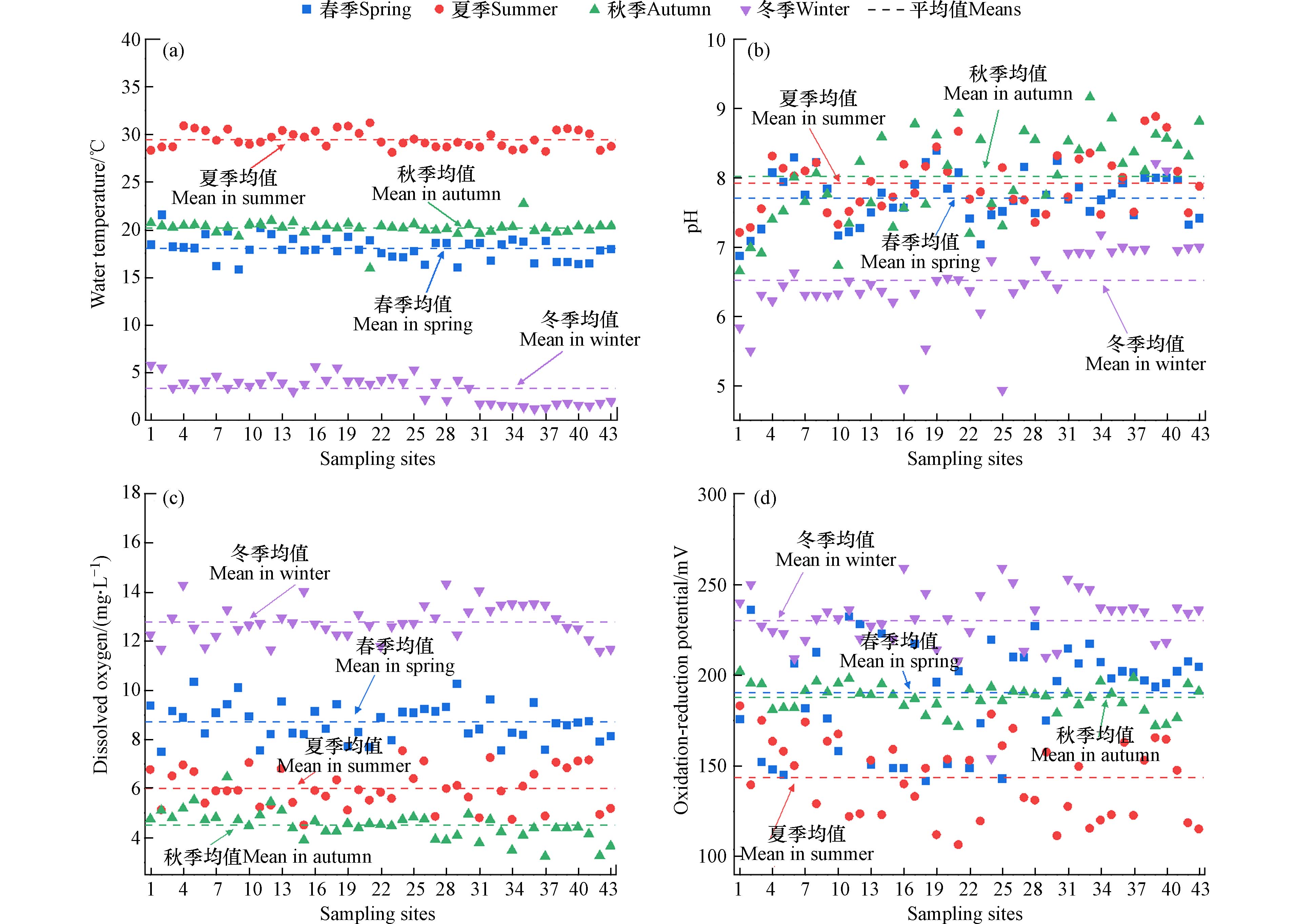
图 2 春、夏、秋、冬四季采样点中水温(a)、pH(b)、溶解氧(c)和氧化还原电位(d)的变化
Figure 2. Variations of water temperature (a), pH values (b), dissolved oxygen (c) and oxidation reduction potential (d) at the sampling points in spring, summer, autumn and winter
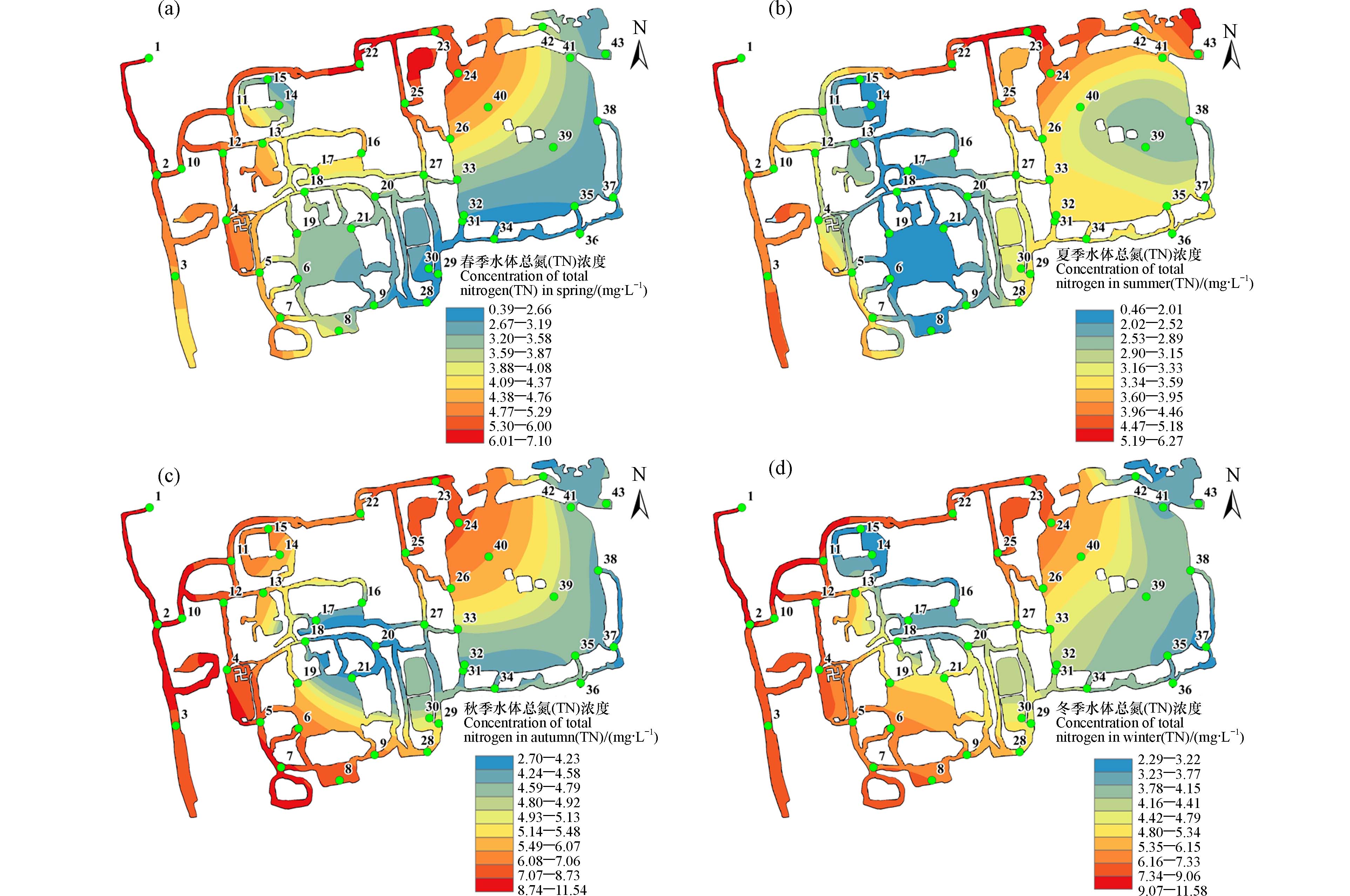
图 3 水中TN浓度在春季(a)、夏季(b)、秋季(c)和冬季(d)的空间分布
Figure 3. The distribution of total nitrogen in water column in spring (a), summer (b), autumn (c) and winter (d)

图 4 水中TP浓度在春季(a)、夏季(b)、秋季(c)和冬季(d)的空间分布
Figure 4. The distribution of total phosphorus in water column in spring (a), summer (b), autumn (c) and winter(d)
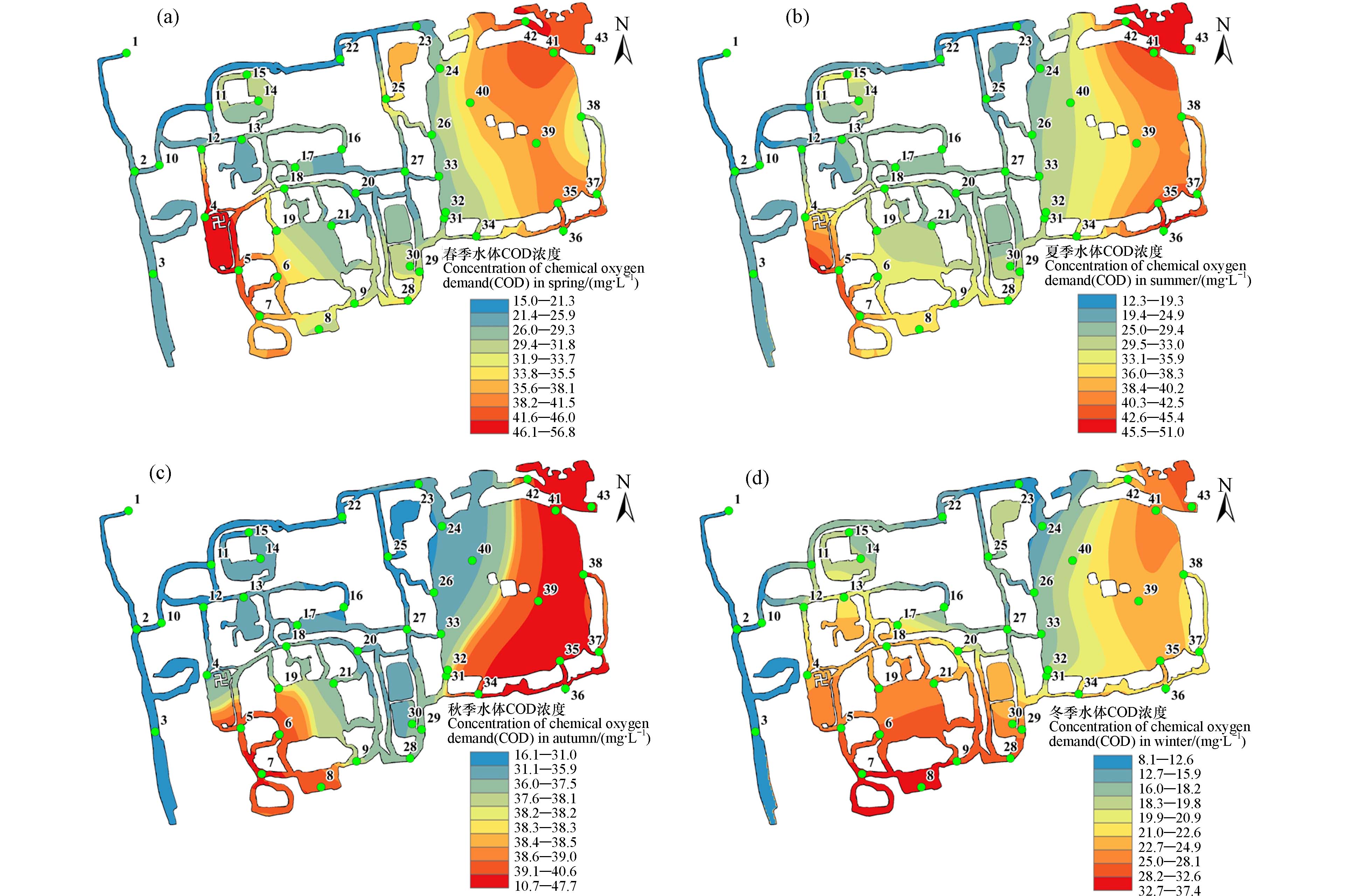
图 5 水中COD浓度在 春季(a)、夏季(b)、秋季(c)和冬季(d)的空间分布
Figure 5. The distribution of COD in water column in spring (a), summer (b), autumn (c) and winter (d)
表 1 主要植物修复区域各季节总磷去除效率
Table 1. Removal efficiency of total phosphorus by season in major phytoremediation areas
修复区域
Phytoremediation area主要植物类型
Main type of aquatic plants总磷去除效率 /%
Removal efficiency of total phosphorus春
Spring夏
Summer秋
Autumn冬
Winter14—15号点 挺水植物 78.3 34.4 72.4 70.9 18—21号点 沉水植物 43.4 6.6 57.2 52.4 28—30号点 挺水植物 63.4 34.6 45.8 65.8  下载: 导出CSV
下载: 导出CSV
-
[1] LIU W, XU Z Q, LONG Y J, et al. Replenishment of urban landscape ponds with reclaimed water: Spatiotemporal variations of water quality and mechanism of algal inhibition with alum sludge[J]. Science of the Total Environment, 2021, 790: 148052. doi: 10.1016/j.scitotenv.2021.148052 [2] BORMANS M, MARŠÁLEK B, JANČULA D. Controlling internal phosphorus loading in lakes by physical methods to reduce cyanobacterial blooms: A review[J]. Aquatic Ecology, 2016, 50(3): 407-422. doi: 10.1007/s10452-015-9564-x [3] LI L X, ZHANG W W, ZHANG M Z, et al. Applications of phytoremediation to treat reclaimed water in urban parks using aquatic macrophytes[J]. Aquatic Ecology, 2022, 56(1): 75-88. doi: 10.1007/s10452-021-09894-x [4] XIAO H, PENG S T, LIU X B, et al. Phytoremediation of nutrients and organic carbon from contaminated water by aquatic macrophytes and the physiological response[J]. Environmental Technology & Innovation, 2021, 21: 101295. [5] ROVIRA A, ALCARAZ C, TROBAJO R. Effects of plant architecture and water velocity on sediment retention by submerged macrophytes[J]. Freshwater Biology, 2016, 61(5): 758-768. doi: 10.1111/fwb.12746 [6] SHAH M, HASHMI H N, ALI A, et al. Performance assessment of aquatic macrophytes for treatment of municipal wastewater[J]. Journal of Environmental Health Science and Engineering, 2014, 12(1): 106. doi: 10.1186/2052-336X-12-106 [7] ZHU Z, LI L X, YU Y N, et al. Distribution, source, risk and phytoremediation of polycyclic aromatic hydrocarbons (PAHs) in typical urban landscape waters recharged by reclaimed water[J]. Journal of Environmental Management, 2023, 330: 117214. doi: 10.1016/j.jenvman.2023.117214 [8] AGHADADASHI V, MOLAEI S, MEHDINIA A, et al. Using GIS, geostatistics and Fuzzy logic to study spatial structure of sedimentary total PAHs and potential eco-risks; An Eastern Persian Gulf case study[J]. Marine Pollution Bulletin, 2019, 149: 110489. doi: 10.1016/j.marpolbul.2019.110489 [9] ZHAO H J, WANG Y, YANG L L, et al. Relationship between phytoplankton and environmental factors in landscape water supplemented with reclaimed water[J]. Ecological Indicators, 2015, 58: 113-121. doi: 10.1016/j.ecolind.2015.03.033 [10] 王文君, 方艳红, 胡晓东. 大渡河河口浮游植物群落时空分布及其与环境因子的关系[J]. 水生态学杂志, 2019, 40(6): 16-23. WANG W J, FANG Y H, HU X D. Spatio-temporal distribution of the phytoplankton community in Dadu River near angu reservoir and its relationship with environmental factors[J]. Journal of Hydroecology, 2019, 40(6): 16-23 (in Chinese).
[11] 陈彦霖, 隋倩雯, 王拓, 等. 温度对序批式膜生物反应器生物脱氮影响及微生物机制解析[J]. 环境工程学报, 2021, 15(1): 65-73. CHEN Y L, SUI Q W, WANG T, et al. Effects of temperature on biological nitrogen removal in batch membrane bioreactor and the microbial community mechanism[J]. Chinese Journal of Environmental Engineering, 2021, 15(1): 65-73 (in Chinese).
[12] SMITH C J, DONG L F, WILSON J, et al. Seasonal variation in denitrification and dissimilatory nitrate reduction to ammonia process rates and corresponding key functional genes along an estuarine nitrate gradient[J]. Frontiers in Microbiology, 2015, 6: 542. [13] GERONIMO F K F, MANIQUIZ-REDILLAS M C, KIM L H. Fate and removal of nutrients in bioretention systems[J]. Desalination and Water Treatment, 2015, 53(11): 3072-3079. doi: 10.1080/19443994.2014.922308 [14] 刘明文, 孙昕, 李鹏飞, 等. 3种水生植物及其组合吸收去除水中氮磷的比较[J]. 环境工程学报, 2021, 15(4): 1289-1298. LIU M W, SUN X, LI P F, et al. Comparison of the absorption and removal of nitrogen and phosphorus from waterbody by three aquatic plants and their combinations[J]. Chinese Journal of Environmental Engineering, 2021, 15(4): 1289-1298 (in Chinese).
[15] LI J H, YANG X Y, WANG Z F, et al. Comparison of four aquatic plant treatment systems for nutrient removal from eutrophied water[J]. Bioresource Technology, 2015, 179: 1-7. doi: 10.1016/j.biortech.2014.11.053 [16] TORRESI E, FOWLER S J, POLESEL F, et al. Biofilm thickness influencesiodiversity in nitrifying MBBRs-implications on micropollutant removal[J]. Environmental Science & Technology, 2016, 50(17): 9279-9288. [17] LONG X E, CHEN C R, XU Z H, et al. Abundance and community structure of ammonia-oxidizing bacteria and Archaea in a temperate forest ecosystem under ten-years elevated CO2[J]. Soil Biology and Biochemistry, 2012, 46: 163-171. doi: 10.1016/j.soilbio.2011.12.013 [18] 曲疆奇, 刘青, 张清靖, 等. 养殖尾水浮床处理系统水生植物根际细菌的氮循环作用机制[J]. 大连海洋大学学报, 2023, 38(1): 12-21. QU J Q, LIU Q, ZHANG Q J, et al. Regulation of bacterial nitrogen cycle of aquatic plants rhizosphere in the ecological floating bed system for aquaculture tailwater treatment[J]. Journal of Dalian Ocean University, 2023, 38(1): 12-21 (in Chinese).
[19] 陈帆帆, 王亚楠, 何圣兵. 不同盐沼湿地类型脱氮除磷效能及影响因素分析[J]. 水处理技术, 2021, 47(1): 95-99,105. CHEN F F, WANG Y N, HE S B. Analysis on efficiency and the influencing factors of nitrogen and phosphorus removal in different salt marsh wetland types[J]. Technology of Water Treatment, 2021, 47(1): 95-99,105 (in Chinese).
[20] ZHU Z, WANG Z F, YU Y N, et al. Occurrence forms and environmental characteristics of phosphorus in water column and sediment of urban waterbodies replenished by reclaimed water[J]. The Science of the Total Environment, 2023, 888: 164069. doi: 10.1016/j.scitotenv.2023.164069 [21] JIANG M Q, ZHOU Y P, WANG N, et al. Allelopathic effects of harmful algal extracts and exudates on biofilms on leaves of Vallisneria natans[J]. Science of the Total Environment, 2019, 655: 823-830. doi: 10.1016/j.scitotenv.2018.11.296 [22] 刘足根, 张萌, 李雄清, 等. 沉水-挺水植物镶嵌组合的水体氮磷去除效果研究[J]. 长江流域资源与环境, 2015, 24(增刊1): 171-181. LIU Z G, ZHANG M, LI X Q, et al. Nitrogen and phosphorus removal of eutrophic water by the mosaic system of submerged-emerged plants[J]. Resources and Environment in the Yangtze Basin, 2015, 24(Sup 1): 171-181 (in Chinese).
[23] SANZ-LÁZARO C, VALDEMARSEN T, HOLMER M. Effects of temperature and organic pollution on nutrient cycling in marine sediments[J]. Biogeosciences, 2015, 12(15): 4565-4575. doi: 10.5194/bg-12-4565-2015 [24] WANG Y Z, LIU M Z, DAI Y, et al. Health and ecotoxicological risk assessment for human and aquatic organism exposure to polycyclic aromatic hydrocarbons in the Baiyangdian Lake[J]. Environmental Science and Pollution Research International, 2021, 28(1): 574-586. doi: 10.1007/s11356-020-10480-1 [25] 王婷, 耿绍波, 常高峰. 人工湿地植物对生活污水净化作用的研究进展[J]. 环境科学与技术, 2013, 36(增刊1): 210-212, 227. WANG T, GENG S B, CHANG G F. Advances in studies on the purification of artificial wetland plants for sanitary sewage[J]. Environmental Science & Technology, 2013, 36(Sup 1): 210-212, 227 (in Chinese).
-
 点击查看大图
点击查看大图
计量
- 文章访问数: 1559
- HTML全文浏览数: 1559
- PDF下载数: 19
- 施引文献: 0
